La croissance des plantes
Dans cette partie, nous verrons que le nombre d'or, la suite de Fibonacci et la croissance des plantes sont très liés.
Angle d’or et divergence d’or
La croissance d'une plante se fait
par les extrémités, dans une zone composée d'une multitude
de cellules indifférenciées qui se divisent puis se spécialisent
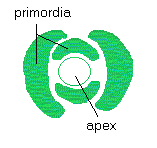
: cette zone est l'apex. Tout autour, de minuscules excroissances se forment,
qui deviendront les futurs organes de la plante (branches, fleurs, feuilles,
...) : ce sont les primordia.
En étudiant la répartition des feuilles autour
de la tige, on constate qu'elle se fait selon trois schémas fondamentaux
: une feuille par nœud , deux feuilles par nœud, trois feuilles
ou plus par nœud (un noeud est une zone d'embranchement, d'où
partent des feuilles, branches ou fleurs).
Parmi les plantes à une feuille
par nœud, il existe les spiralées : les feuilles apparaissent
une à une le long de la tige en formant des spirales. Sur ces plantes,
chaque primordium s’écarte de l’apex et, finalement, se
transforme en une feuille ou un pétale. La spirale engendrée
par les primordia dans l’ordre de leur apparition est appelée
spirale génératrice, les primordia les plus anciens étant
les plus éloignés du centre.
En 1837, Auguste Bravais et son frère Louis, observèrent un détail essentiel. Ils tracèrent les demi-droites qui joignent le centre de l’apex au centre des primordia et mesurèrent les angles séparant deux primordia successifs vus de l’apex. Les angles entre les centres de deux primordia approchent les 137°10’, c'est à dire la divergence d'or.
Cette image représente la répartition des primordias en partant de l'apex : celle du milieu est tracée avec une divergence (angle entre deux primordia) égale à la divergence d'or ; les deux autres en diffèrent de moins d'un degré.

Si
en regardant la tige d’une spiralée par le côté
de l'apex, on observe la disposition des feuilles, on remarque qu’après
un certain nombre de tours parcourus le long de la spirale génératrice,
une feuille se retrouve exactement en dessous de la première feuille
que l’on observe. On peut alors :
- compter le nombre de feuilles situées après la première
(c’est la n° 0, donc on ne la compte pas) jusqu’à parvenir
à la feuille qui est exactement alignée avec la première
(n° x).
- compter le nombre de tours que fait la spirale génératrice
autour de la tige entre ces deux feuilles de référence (y).
Toute les plantes à phyllotaxie de type « spirale » ont
des fractions x / y égales à 2 / 1 ou 3 / 1 ou 5 / 2 ou 13 /
5 ou 21 / 8 ou 34 / 13, etc. Les numérateurs et les dénominateurs
de toutes ces fractions appartiennent à la suite de Fibonacci.
Exemple de quelques répartitions : 
Explication de la présence du nombre d’or chez ces plantes
La présence du nombre d’or dans la phyllotaxie des plantes a suscité de nombreuses investigations. La première explication a un lien avec l’ensoleillement : «Si les feuilles (et par conséquent les rameaux) d’une plante étaient espacées sur la tige par des intervalles d’exactement 137°30’ 28’’, aucune feuille ne se situerait exactement au-dessus d’une autre, ce qui diminuerait l’ombre portée par cette feuille sur les autres situées plus bas» (Bell, 1993, p222). Il y a donc là une première règle d‘optimisation de la collecte des photons, optimisation qui pourrait s’être mise en place au cours de l’évolution.
Mais pourquoi l’angle de divergence est il égal
à l'angle d’or ? Pour le découvrir, il faut revenir au
début de la croissance de la tige, lorsqu’elle n’était
qu’un bourgeon. Au cours de la croissance, l’apex croit en gardant
la même forme, mais les primordia ne croissent pas aussi vite et s’éloignent
donc de l’apex ; c’est dans cet espace, entre les primordia et
l’apex, que de nouveaux primordia apparaissent. Cependant ceux-ci gardent
tout au long de la croissance la disposition qu’ils avaient initialement.
Les règles d’apparition des primordia sont partagées entre
deux hypothèses :
- Le critère de W. Hofmeister, selon lequel les primordia apparaissent à intervalle régulier dans le plus grand espace disponible laissé par les primordia précédents.
- Le critère de Marie et Robert Snow, selon lequel
un nouveau primordium apparaît dès qu'il s'est formé un
espace libre de taille suffisante. Ce critère a l'avantage d'expliquer
le mode spiralé et le mode verticillé.
Prenons le cas d'une plante verticillée où les feuilles poussent
par paires opposées, l'apex ressemble alors à ceci :
Lors de la croissance, selon le critère Snow, on obtient :

D'où la formation de paires de feuilles. Mais si on incise l'apex, il se produit un décalage dans la disposition et comme l'a expérimenté le couple Snow, on obtient une plante spiralée :
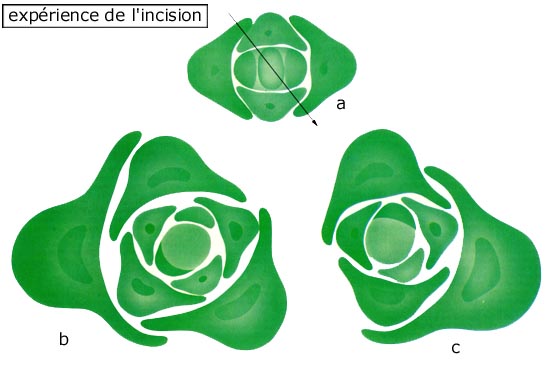
Pour expliquer
comment chaque primordium connaît quelle est la plus grande place disponible
ou quand la place est de taille suffisante, le mathématicien Alan Turing
a émis l'hypothèse que chaque primordium émet autour
de lui une substance inhibitrice empêchant la formation d'autres primordia
à sa périphérie. En simulant numériquement l'apparition
des primordia à partir du critère d'Hofmeister, Stéphane
Douady et Yves Coudier ont mis en évidence que l'angle de divergence
dépendait fortement d'un facteur appelé G.
G = vT/Ro
Où :
v est la vitesse d'éloignement des primordia par rapport à l'apex
T est la périodicité d'apparition des primordia
Ro le rayon où elles apparaissent.
En résumé lorsque la valeur de G est grande, les primordia s'éloignent
rapidement de l'apex et/ou le temps d'apparition entre deux primordia est
long :
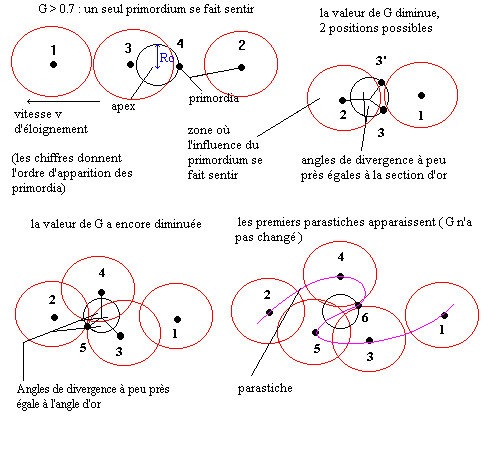
Lorsque G baisse jusqu'à la valeur 0.7, chaque nouveau primordium ressent l'influence de deux primordia, et vient former un angle qui se rapproche de l'angle d'or. Donc à chaque fois que G a diminué au point que chaque nouveau primordium ressente l'influence d'un primordium supplémentaire, l'angle de divergence tend un peu plus vers le nombre d'or, ceci s'appelle une brisure de symétrie.
Lors de la première brisure, le primordium peut apparaître
à deux endroits, mais une fois apparu, la position choisie forcera
les primordia suivants à apparaître selon une spirale tournant
à droite ou à gauche. D'ailleurs dans la nature on trouve autant
de plantes dont la spirale génératrice (spirale reliant les
feuilles d'une tige dans leur ordre d'apparition) tourne vers la droite que
vers la gauche. Normalement, il existe à chaque brisure de symétrie
un deuxième angle de divergence possible mais mis à part pour
la première brisure de symétrie (cf. ci-dessus), seul l'angle
qui tend vers la section d'or est possible si G diminue progressivement.
Par contre, si G diminue brutalement, les autres changements d'angle de divergence
sont possibles. Or, chez les plantes, le facteur G diminue progressivement
au cours de la croissance : il est fort lorsqu'elles germent puis il décroît
lorsque les premières feuilles apparaissent, et la floraison s'accompagne
également de la diminution de la valeur de G. C'est pourquoi l'angle
de divergence tend vers l'angle d'or.
Parastiches, pétales et suite de Fibonacci
De nombreux végétaux présentent des spirales apparentes à l’œil nu, comme les hélice formées par les écailles d’un ananas ou d’une pomme de pin ou les aiguilles d’un cactus. Ces spirales sont appelées parastiches. Il existe deux types de parastiches : celles qui tournent dans le sens direct et celles qui tournent dans le sens indirect (sens des aiguilles d’une montre). S’il on compte le nombre de parastiches tournant dans un sens et dans l’autre, on trouve deux nombres consécutifs de la suite de Fibonacci ; de plus l’étude de ces spirales a permis de conclure qu’elles étaient logarithmiques.

Ainsi, pour l’ananas, on compte 8 parastiches dans le sens indirect
et 13 dans le sens direct, pour la pomme de pin 5 et 8 et selon les variétés
de tournesols, 21 et 34, 34 et 55, 55 et 89, voir 89 et 144. On trouve rarement
d’autres nombres,les exceptions les plus notables les doubles de ces
nombres.
Souvent, chez les fleurs dont les pétales sont disposés en hélice
autour du cœur on trouve des nombres de Fibonacci. Les lys ont 3 pétales,
les boutons d’or 5, la plupart des soucis 13, les asters 21.
Le nombre d’or est lié à une spirale
logarithmique. On sait qu’une spirale logarithmique (ou hélice)
peut s’étendre indéfiniment vers l’extérieur
ou vers l’intérieur. Elle garde donc toujours la même forme,
indépendamment de sa dimension. La forme est conservée, quelle
que soit la taille.
On voit donc que lorsque les éléments tous semblables d’une
structure modulaire, ici les primordia, les pétales,..., grandissent
sans changer de forme, les restructurations permanentes qu’ils subissent
pour minimiser les contraintes de compression tendent à les organiser
le long de spirales logarithmiques. Le calcul du nombre de spirales dans un
sens puis dans l’autre mène à des couples de nombres entiers
qui font partie des séries de Fibonacci.
Par exemple, dans le cas de la marguerite, les bourgeons floraux ont tous
la même forme (ce sont de simple tubes de section circulaire), mais
pas la même taille. Ils s’auto organisent au fur et à mesure
de leur croissance selon des spirales logarithmique centrifuges. On peut faire
alors les mêmes observations sur une pomme de pin, les éléments
modulaires changent de taille sans changer de forme.
Conclusion
On peut donc conclure que la croissance et l'organisation des plantes sont bâties sur des principes mathématiques (suite de Fibonacci, nombre d'or ) qui sont l'expression optimale des gènes de la plante.
![]()
![]()