ARBORESCENCES
Economie de travail et angles de branchement
Parmi les diverses méthodes permettant de relier plusieurs points à un point central, les arborescences sont les plus économiques. Partant du principe qu'entre deux solutions, pour résoudre un problème, l'évolution retient toujours la plus économe en terme de matériaux et d'énergie, on peut s'attendre à ce que les "arborescences" soient très fréquemment représentées dans les constructions naturelles. C'est effectivement le cas et, que ce soit dans le monde végétal ou dans le monde animal, les exemples abondent : l'irrigation des poumons, des reins mais surtout les branches des arbres.
Un simple coup d'oeil à ces structures montre une parenté de formes qui intrigue. En effet, que peut-il y avoir de commun entre l'arbre et l'arborescence d'artères rénales ? Quelles sont la ou les lois sous-jacentes qui peuvent expliquer cette parenté de forme dans des contextes si différents. Le but de cet article est de vous montrer si toutes les arborescences partagent des règles communes ou si, malgré une ressemblance qui pourrait n'être que superficielle, chaque type d'arborescence est unique et dépend de l'organisme considéré.
Cet article est
consacré à la mise en évidence de la loi de Roux & Murray, loi simple qui
montre qu'à chaque bifurcation, le rapport du diamètre des branches et l'angle
qu'elles font entre elles sont liés. Plutôt que de commencer par décrire la loi
de Murray, nous allons procéder d'abord
par une série de mesures dans la nature, puis traiter et reporter ces mesures
sur un graphe et enfin nous analyserons les propriétés du graphe.
MESURES
Il existe deux cas de figures possibles : une bifurcation
simple avec des branches alternes ou une bifurcation avec des branches
opposées. Nous étudierons surtout les variantes du premier cas, car le second
est en fait un cas particulier du 1er.
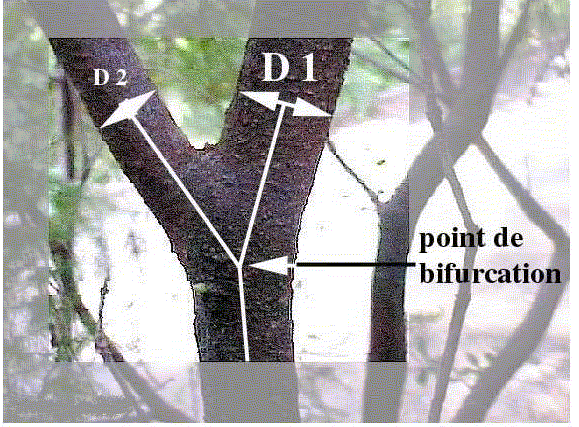
Au niveau d'une fourche, effectuons deux types de mesures :
les valeurs des diamètres D1 de la "grosse branche" G et D2 la "petite
branche" P et l'angle A et l'angle B en degrés, A étant l'angle entre
les deux branches secondaires et B celui entre D1 et la droite passant par
le tronc ou la branche maîtresse.
A partir des mesures, nous pouvons calculer l'angle A en
fonction du diamètre D2/D1 (ces valeurs traduisent le resserrement progressif
de l'angle que fait la branche secondaire par rapport à la branche principale
lorsque la branche secondaire augmente en diamètre) et l'angle (90-B) ° en
fonction du diamètre D2/D1 (ces valeurs traduisent la déflexion de plus en plus
forte de la branche principale lorsque la branche secondaire augmente en
diamètre). À partir d’une bonne quantité de mesure, le savant Murray a obtenu
un graphe très intéressant.
INTERPRÊTATION DES DONNÉES : LES DIAGRAMMES DE MURRAY
Les diagrammes de Murray, qui expriment la relation entre le
diamètre des branches et l'angle qu'elles font entre elles, sont constitués
par trois courbes, dénommées I, II et III (adapté de Stevens P.S., 1978. Les
formes dans la nature, Seuil, 232 pages).

On utilise la courbe I pour l'angle A que fait la branche secondaire
par rapport à la branche principale, la courbe II pour l'angle (90-B)° de
déflexion de la branche principale et la courbe III pour l'angle C entre l'une
quelconque de deux branches opposées et la branche principale.

Remarquons que lorsque deux branches égales, opposées l'une à
l'autre, se séparent du tronc, il n'y a pas déflexion de celui-ci, alors qu'une
seule branche secondaire qui se sépare de la branche principale la défléchit
d'un angle égal à (90-B) ° ; tout ceci est en accord avec l'observation de
Léonard de Vinci :
"On
trouve les branches des plantes suivant deux positions différentes : soit
opposées l'une à l'autre, soit non opposées. Lorsqu'elles sont opposées, la
tige centrale n'est pas courbée ; si elles ne sont pas opposées, la tige
centrale est courbée."
Sur le graphe de Murray nous voyons que la courbe I qui
représente l'angle A que fait une branche secondaire avec la branche principale
est relativement plate, c'est à dire que l'angle entre deux branches adjacentes
varie relativement peu, même pour des branches de tailles différentes. Si la
branche principale se sépare en deux branches égales, avec un rapport des
diamètres égal à 1, ces branches divergent d'environ 75°, parfois 70°. Si la
bifurcation engendre de petites branches - avec un rapport du diamètre de la
branche secondaire à celui de la branche principale tendant vers 0, la
divergence est voisine de 90°. Les angles de branchement varient donc à
l'intérieur d'une plage étroite allant de 90° à 70°.
ATTENTION : les diagrammes de Murray ont été établis pour des
arborescences d'écoulement telles que les artères et les veines. Les
mêmes courbes pour le végétal ont des aspects légèrement différents. En
particulier la courbe I est beaucoup moins "plate" et descend jusqu'à
des angles de 20° ou 30° lorsque les diamètres des branches secondaire et
principale sont identiques (rapport D2/D1 = proche de 1). Les angles de
branchement (angle A) varient donc dans une plage beaucoup plus large que pour
les artères et les veines, et il y a une nette tendance au resserrement des
angles lorsque les diamètres sont voisins ou égaux.
Par contre la variation de l'angle (90-B)° (courbe II),
représentant l'augmentation de la déflexion de la branche principale en fonction
de l'augmentation de la taille de la branche secondaire, varie de 90° pour les
petites branches (elles ne provoquent pas de déflexion), à 45° (déflexion
maximale). En effet, pour éviter des ruptures dues au poids, il est nécessaire
que la branche principale s'infléchisse de manière à ramener le plus possible
le poids de la branche secondaire vers le centre de gravité.
SIGNIFICATION BIOLOGIQUE DES DIAGRAMMES DE MURRAY
La relation entre le diamètre des branches et l'angle
qu'elles font entre elles obéit à une loi d'optimisation des écoulements. Ici
l'écoulement correspond au transport de la sève dans l'arbre. Il pourrait
s'agir d'autres types d'écoulements, par exemple celui du sang, et la loi de
Murray a été précisément démontrée dans ce contexte. C'est donc une loi
universelle (avec des variantes selon le type d'organisme), qui s'applique à
tous les écoulements canalisés par des tuyaux creux dans lesquels circule un
liquide dont le trajet doit être le plus court possible et subir le moins de
ralentissement possible.

En résumé.
Lorsque la branche (ou l'artère) principale est beaucoup plus large que la
branche (artère) secondaire, la sève (sang) fournit moins de travail en
s'écoulant principalement dans la branche (artère) la plus large et en
minimisant le trajet parcouru dans la branche (artère) la moins large. Par
contre, si les deux branches (artères) ont sensiblement le même diamètre, la
sève (sang) pourra s'écouler dans la branche (artère) secondaire sans
augmentation sensible de l'effort et l'angle de branchement oscillera entre 60
et 75° dans le cas des systèmes artériels et veineux et entre 60 et 30° dans le
cas des arbres.
Nous pouvons
énoncer ainsi deux règles générales :
1) Plus la branche secondaire est petite par rapport à la
branche principale, plus elle s'éloigne de celle-ci à un angle voisin de 90°.
2) Plus le diamètre de la branche
secondaire est proche de celui de la principale, plus celle-ci s'infléchit.
Pourquoi les petites branches
partent-elles à 90° par rapport aux branches principales ?
Soit un liquide circulant de A vers D et un point P qui doit
être relié au réseau (P pourrait être une feuille pour un arbre). L'alternative
est la suivante :

à soit
le liquide circule de A à C dans la branche principale puis emprunte une
branche courte pour aller de C à P
à soit
le liquide circule directement par une branche secondaire BP qui est du même
diamètre que CP, mais plus courte.
La loi d'économie suggère que le trajet direct BP sera
privilégié par rapport au trajet BCP. Cependant, si l'on considère les vitesses
d'écoulement, le faible diamètre de la section BP va entraîner une forte
résistance à l'écoulement, qui va ralentir le flux. Par contre, dans la branche
principale, le flux est rapide jusqu'au point C, puis ralenti entre C et P,
mais la distance CP est très courte.
Pour que le trajet BP soit réellement plus rapide que le
trajet BCP, il faut :
à que
la bifurcation au point B se fasse avec un angle aussi fermé que possible,
à que
le diamètre de la branche secondaire BP tende à devenir aussi gros que le
diamètre de la section BC, de manière à ce que l'écoulement ne soit pas ralenti
par les effets de frottement.
Si ces conditions ne sont pas remplies, alors le trajet BCP
sera le plus efficace.
FROTTEMENTS ET VISCOSITÉ
Pourquoi faut-il que le trajet dans les branches de petit diamètre
soit le plus court possible ? Ceci est dû aux forces de frottement contre
la paroi du tube :

On voit que le diamètre "utile" pour la circulation
du fluide est plus petit que le diamètre réelle. La
différence entre le diamètre "utile" et le diamètre
"réelle" correspnd à la zone de frottement
le long des parois.
Ceci explique par ailleurs pourquoi dans un arbre la somme
des diamètres réels des branches principales est supérieure au diamètre du
tronc (de 2 à 2,5 fois plus) :
Mais si l'on tient compte seulement du diamètre
"utile" à l'écoulement, alors la somme des diamètres des tuyaux
d'écoulement dans les branches est sensiblement égale à la somme des diamètres
des tuyaux dans le tronc.
PESANTEUR VS. ENCOMBREMENT
Si les lois de Roux & Murray s'appliquent bien à tous les
réseaux distribuant des liquides, il existe des différences entre ce que l'on
observera localement, selon le contexte physique dans lequel se développe
l'organisme vivant.
Ainsi, les angles de bifurcations entre deux branches de même
diamètre sont en moyenne plus fermés pour les arbres que pour les réseaux
d'artères et de veines.
Ceci est dû au fait que les arbres poussent en s'opposant à
la pesanteur, mais ne manquent pas d'espace pour s'étaler. La gravité tend à
écarter les branches et à les ramener vers le sol, d'où la nécessité d'avoir
des angles aussi fermés que possible pour que les branches ne s'écartent pas
trop du tronc et soient moins exposées à la pesanteur tout en offrant un
maximum de surfaces exposées à la lumière (les feuilles).
Dans les réseaux d'artères et de veines, la pesanteur est
secondaire par rapport à la lutte pour l'espace disponible, celui-ci étant très
limité par définition. Les angles pourront donc être plus ouverts, sans
atteindre cependant des angles trop élevés car il se produirait alors un net
ralentissement du flux sanguin.
ARBORESCENCES D'ECOULEMENT vs. ARBORESCENCES DE SUPPORT
Nous avons dit au début de ce chapitre que les arborescences
telles que celles de la Gorgone (Cnidaire) Paramuricea
chamaleon sont profondément différentes de celles
vues dans le tableau ci-dessus. En effet, ils ne
s'agit pas comme dans les cas précédents d'arborescences d'écoulement, mais d'arborescences
de support. En effet, les polypes de la Gorgone se disposent les uns par
rapport aux autres pour optimiser la collecte des micro-éléments planctoniques
qu'ils capturent avec leurs tentacules (micro crustacés et larves en
particulier). Le squelette souple secrété par chaque polype permet à toute la
colonie de se déployer dans le courant, perpendiculairement à celui-ci. Le
squelette est donc principalement une structure de support et, à la limite,
chaque polype pourrait être complètement autonome par rapport à ses voisins.
C'est d'ailleurs ce qui se passe chez certaines formes coloniales de Cnidaires
où les polypes individuels se disposent les uns à côté des autres sur des axes
de Gorgones mortes (ou sur des éponges arborescentes) : le "Mimosa de
mer", Parazoanthus axinellae
est l'une de ces espèces d' Anémones qui s'installent
sur des supports variés, ce qui permet aux polypes de bien s'étaler dans le
courant.
CONCLUSION
Le but de cet article a été de montrer, pour les réseaux
arborescents d'écoulement, que quelques lois simples, universelles dans leur
application, rendent compte d'une bonne part des organisations observées.
Trajets minimaux et économie d'énergie contrôlent ainsi de façon rigoureuse la
morphogenèse de tels réseaux d'écoulement de fluides. Les arborescences qui
n'ont pas pour rôle de distribuer un fluide ont d'autres règles de
construction, c'est le cas de la disposition des feuilles chez une plante, où,
si vous ne l'avez pas encore lu, l'optimisation de la surface d'ensoleillement
fait appel à un nombre extraordinaire, le nombre d'or.
![]()